

Journal of Creation 34(1):79–85, April 2020
Browse our latest digital issue Subscribe
Out of Babel—not Africa: genetic evidence for a biblical model of human origins
The recent human evolution story is a quagmire of ever-changing speculations that vary and conflict in their dates and geographical specifics with each new paleoanthropological or archaic DNA sequence discovery. The current general consensus is often termed the recent out-of-Africa model that postulates anatomically modern humans migrated out of Africa about 100,000 to 200,000 years ago. While various versions of the hypothetical evolutionary story exist, an increasing body of research in both the creationist and secular spheres is vindicating the Bible’s timeline of a global-Flood-based genetic bottleneck about 4,500 years ago and a recent creation about 6,000 years ago. In this review, I cover the overwhelming evidence from genetics research of various flavors that fully vindicates a biblical picture of human origins.

During the 1980s and early 1990s, much of secular paleoanthropology was equally divided on how modern humans originated between one idea known as multiregionalism and the other known as Recent African Origins (RAO). The multiregionalism model proposed that humans evolved in different parts of the world simultaneously, with various genes being spread through intermixing and interbreeding. The RAO proposed that humans first evolved in Africa and then dispersed out from there across the world. During the 1990s, especially after the publication of the African Eve hypothesis, the RAO began to rapidly pervade academic circles and has since become the dominant model along with many twists and variants to its central premise.1
The fossil record for human evolution is filled with gaps. Thus, inconsistencies and biogeographical confusion abound and scientists have begun to look to the emerging field of DNA analysis. Starting with the mitochondrial DNA work of Cann, Stoneking, and Wilson in 1987 and steadily progressing from there using more advanced technologies, the RAO model began to pick up speed and eventually dominated the academic landscape. The general idea is that mitochondrial and nuclear DNA diversity is greatest in Africa, and, based on known human diversity along with a whole host of other questionable genetic assumptions, which are based in turn on a neutral model of evolution (see Carter 2009),2 humans evolved in Africa about 100,000 to 200,000 years ago, with specific dates and out-of-Africa migration events depending on the model theorist(s).
Just as the RAO seemed to reach its peak of acceptance, the emerging field of ancient DNA sequencing revealed that admixture between anatomically modern humans and Neandertals and Denisovans had taken place. This was quite surprising because: 1) Neandertals were not supposed to be part of the African origins, and so were not supposed to be Homo sapiens; and 2) Denisovans were completely unknown. RAO sub-models then quickly began emerging to account for this inconvenient data. According to recent reviews, four main human origins models now exist and compete with one another:
- RAO (classic): Modern humans first developed in Africa about 100,000 years ago and then migrated throughout the world. Existing archaic human populations (i.e. Neandertal and/or Homo erectus) in various areas of the world were then replaced by the RAO migrating populations, with little to no hybridization between the populations.
- RAO With Hybridization: This model is similar to the classic RAO above, but includes a greater level of hybridization taking place between the migrating populations and the indigenous archaic populations being encountered and displaced.
- RAO Assimilation: This model, like the other two above, accepts a recent African origin for modern humans. Unlike the previous two models, it includes replacement and/or extensive migration of populations as the major driving factor in the emergence of modern humans. This model focuses on the importance of pervasive gene flow and population admixture in conjunction with changing environmental conditions.
- Multiregionalism: This model denies a recent African origin for modern humans, based on biogeographical data that continue to emerge from paleontology and archaeology. This model also promotes the role of broadscale genetic continuity over time and gene flow between populations. The basic premise is that modern humans arose not only in Africa, but also in Europe and Asia from their H. erectus type Pleistocene ancestors.
If this quagmire of competing models was not confusing enough, researchers are at odds concerning the origins of modern human within Africa before the alleged global dispersion(s) ever occurred. As noted by Henn et al. in 2018, there are four of these submodels competing with each other.1 The first is known as African multiregionalism, which maintains that there were multiple points of modern human origins across Africa, with continuing admixture between populations. The second proposes that there was a single point of origin, with range expansions and local extinctions, but with one line predominating. The third proposes that there was a single point of origin, with range expansion and one line predominating, but slightly lesser-evolved populations breaking off shortly before the dispersion, and occasional admixing. The fourth proposes that there was one point of modern human origins, with multiple lines of archaic humans evolving side by side with a small amount of admixture occurring shortly before the dispersal.
Archaic DNA sequencing deepens the RAO quagmire
The field of archaic human DNA sequencing, along with massive-scale sequencing of numerous genomes of modern humans worldwide, has completely revolutionized the secular field of human evolution. In these studies, the nuclear genomes of multiple archaic Neandertals and Denisovans have been sequenced at various levels of completeness. In addition, an even larger number of mitochondrial DNA sequences have been obtained. Instead of confirming evolutionary predictions, especially those associated with the RAO model, the results have brought more questions than answers.3
Neandertals are known by both archaeological and paleontological findings and are believed to have lived in the Middle East, Europe, and various regions in Asia. In contrast to the rich amount of evidence for Neandertals, Denisovans are only known from ancient DNA extracted from a single finger bone and three teeth and are thought to have inhabited parts of East and South-East Asia. Anatomically modern humans are thought to have existed about 300,000 years ago, based on the recent fossil finding in North Africa at Jebel Irhoud.4,5 However, Neandertals are believed to have lived from 450,000 years ago up until only 40,000 years ago.3 Thus, modern humans and their archaic cousins are believed to have overlapped both in time and geography.
Not surprisingly, archaic human DNA is nearly identical to that of modern humans, although its sequence quality can often be questionable.6 According to the consensus creationist model, Neandertals are nothing more than people groups of post-Flood humans. However, evolutionists are in a constant quandary over their interbreeding (admixture) with anatomically modern humans, despite the fact that so-called archaic traits like sloping foreheads and pronounced brow ridges can still be found among modern humans today. Along these lines, the ever-increasing timeline of admixture is causing confusion for the RAO.
The first major discrepancy for RAO that came from archaic DNA studies arose when it was discovered that modern non-African humans shared higher levels of genetic ancestry with archaic humans than they did with African populations.7 Making matters worse, a 2017 study of mitochondrial DNA showed that anatomically modern humans were admixing at least 270,000 years ago, pushing the date beyond the outer limit of about 200,000 years for RAO.8 But the evolutionary confusion for RAO has now gotten much worse. In 2019, an extensive genomic analysis of archaic and modern human genomes has shown that large segments of Neandertal chromosomes span across centromeres, where recombination does not occur. These segments have also been found to span across 21 different genic regions, yielding a 600,000-year timeline to the last common ancestor between humans and Neandertals.9 This new DNA study, using the evolutionists’ own system of reckoning, triples the RAO maximum.
And now, a recent 2020 study has thrown yet another wrench into the evolutionary machine of the RAO paradigm which shows that genomic regions of Neandertal ancestry are present in individuals of African descent at much higher levels than previously believed.10 The RAO advocates have attempted to explain this anomaly by claiming that humans with Neandertal ancestors migrated back down into Africa from Europe on multiple occasions, spreading their genes among African populations.
Mitochondrial genetic clocks refute RAO
The evolutionary molecular genetic clock has a long and nefarious history in the annals of modern biology (reviewed by Tomkins and Bergman in 2015).11 The general paradigm of an evolutionary genetic clock typically uses an alignment of multiple DNA sequences, along with a variety of statistical models to provide rates of hypothetical evolution—often between unrelated taxa that most creationists would deem separate created kinds. Standard genetic clock studies routinely use deep-time calibrations derived from paleontology and assume a paradigm of macroevolution. And quite notably, clock-derived divergence dates commonly conflict with those taken from paleontology, despite the fact that deep-time calibrations are built into the algorithms. This is consistently true with analyses of human evolution that also add further bias by using chimpanzees as an evolutionary ancestor outgroup.2 Needless to say, this type of approach is not empirically based.
So what kind of genetic clock data could be achieved if the evolutionary assumptions were not used and an empirical approach to the analysis was taken? As it turns out, research by both secular and creation scientists indicates a date of modern human origins of not more than 5,000 to 10,000 years (reviewed by Tomkins in 2015).12 For the purposes of this current review, I will briefly summarize these earlier findings before describing more recent efforts that confirm and refine them.
Evolutionists first noticed a problem with their paradigm in 1997 when human mitochondrial DNA mutation rates were first being analyzed.13 As stated in the original paper: “Using our empirical rate to calibrate the mtDNA molecular clock would result in an age of the mtDNA MRCA [most recent common ancestor or the first human woman] of only ~6,500 years.” This study was ridiculed and maligned, not for the methods used, but for the young age of the non-evolutionary conclusions that were reached.14 In 2012 and 2013, two major secular papers were published that each analyzed the rare mutational variants in human protein coding exons among ~9,000 individuals of various ethnicities and found that, based on demographic models, as opposed to standard evolution-calibrated models, the rare mutational variation in the human genome could be no older than 5,000 to 10,000 years.15,16
While these secular studies were extremely revealing, and directly opposed the timeline claims of RAO, creationist researchers, unhindered by constraints of political academic correctness, began to refine the model further. In 2013, Jeanson demonstrated that the empirical mitochondrial DNA mutation rate in round worms, fruit flies, water fleas, and humans gave a maximum creation age for each of them of not more than 10,000 years ago.17 Jeanson’s results for humans lined up well with the research of Sanford and Carter, who statistically compared over 800 mitochondrial DNA sequences and reconstructed a close approximation of ancestral Eve’s original mitochondrial genome.18,19 They showed that, on average, any given human is only about 22 mutations different than the ancestral Eve sequence, although some may be as many as 100 mutations different. Based on an empirical estimate of human mitochondrial DNA mutation of about 0.5 per generation, as reported in the literature, they estimated that 200 generations would be needed to accumulate 100 mutations—a time of less than 6,000 years. In 2018, Carter, Lee, and Sanford further refined the mitochondrial DNA Eve consensus sequence and even developed haplogroups.20 A haplogroup is a group of genomic variations that are inherited together from a single parent. This work further vindicated a single ancestral genome and also helped to provide some clarity on the dispersion of post-Flood people groups originating in the Middle East, in contradiction to the claims of RAO.
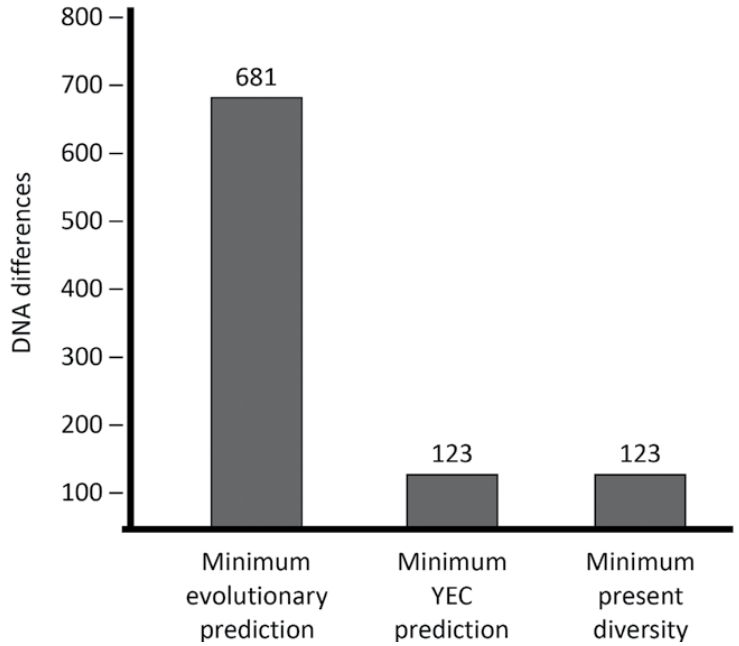
Outside of the reconstructive work by Carter for the ancestral mitochondrial Eve genome, most early mutation rate studies had only been done with what is known as the D-Loop region which represents a mere 7% of the total sequence (16,569 base pairs).21 Also, these analyses had only been done in non-Africans. It is the evolutionary belief that an increased amount of mitochondrial DNA diversity in Africans is indirect evidence for the RAO.2 While these earlier studies gave strong indication of a young-earth timescale, more work needed to be done to confirm the original findings. In 2015, Jeanson utilized newly available data sets of complete mitochondrial DNA genomes and showed that the mutation rate in European individuals fully matched with a 6,000 year timescale and fully captured the mitochondrial DNA diversity in non-African people groups.22 His results included the major Eurasian people groups and representative individuals of native people groups in the Americas. Most importantly, his predictions used a mutation rate based on the entire mitochondrial genome that not only affirmed the previous rate derived from just using the D-Loop, but soundly refuted the evolutionary RAO timeline (figure 1).22
But what about the diversity among Africans? In a follow-up study, Jeanson investigated the global mitochondrial DNA diversity further, incorporating African and Asian data and also included improved information on generation time based on demographics.22 Surprisingly, he found that African females, on average, married much earlier in life (average of 15 to 19 years) than non-Africans, which would increase generation time and thus diversity. And interestingly, the highest divergence was not found among Africans but between an African San individual and an Asian Taiwanese Aborigine.23 This makes perfect sense because Africans and Asians would have split after the Flood due to the events at the Tower of Babel, from which most ethnic groups would owe their origins. Indeed, Jeanson showed how major mitochondrial DNA lineages followed a maternal ancestry back to the three wives of Noah’s sons (figure 2).22
The newer 2016 study further clarified this ancestry with the addition of African and Asian data, and also showed how a split leading to the Asian and African lineages post-Flood may have actually occurred before the Flood. One has to keep in mind also the mechanism of pre-Flood ancestries in the different lineages of Noah’s sons’ wives as well. Nevertheless, the new, more comprehensive data confirmed the material ancestry of three main lineages connected to the three wives of Noah’s sons. And finally, the new data also confirmed and extended the earlier results showing the initial estimates of mutation rate based on non-Africans are not only still consistent with a biblical timeline but can be used to predict mitochondrial DNA diversity whether African or non-African.

Y-chromosome clocks refute RAO
While the mitochondrial data has spectacularly confirmed the biblical timeline for the maternal human lineage, what about the data on the paternal side? As it turns out, Y-chromosome analyses are just as revealing. In addition to their early creationist work on mitochondrial DNA, Sanford and Carter also analyzed Y-chromosome diversity among modern humans and found there to be only about 300 mutations, on average, that were different from the consensus sequence for a Y chromosome representing the original Adam.19 In fact they found that if a normal mutation rate for the Y chromosome of about 1 mutation per generation was assumed, that only about 300 generations corresponding to about six thousand years would be needed to get 300 mutations worth of modern human Y-chromosome diversity.
In 2018, Carter, Lee, and Sanford further refined the history of the Y-chromosome sequence along with similar analyses for the mitochondrial genome.20 Quite notably, they discovered that both the maternal and paternal unrooted phylogenetic analyses gave very similar results. The first key similarity observed was that the branching pattern displayed in both DNA datasets indicated a large expansion of human people groups, with many new branches being formed from closely related ancestors. The second noteworthy trend was that for both chromosomes, mutation rates along various branches were not the same over time, yet when averaged out, both genomes were less than 10,000 years old. Another interesting outcome from the study was that both phylogenetic trees revealed a starburst pattern centering around specific historical individuals derived from the Middle East. All of the data combined showed that, with a high degree of confidence, the actual sequences of historical individuals that gave rise to each branch in both family trees could be determined, including the ancestral Eve and Adam mitochondrial and Y chromosomes, respectively (within a reasonable level of confidence).
To help further resolve the paradigm of a Y-chromosome genetic clock, in 2019 Jeanson and Holland downloaded newly available sequences for the Y chromosome that were derived from individuals in large human pedigrees. The data were considerably more comprehensive and contiguous thanks to new long-read DNA sequencing technologies.24 The Y chromosome is particularly useful in studying human pedigrees, common variants, and mutations, because it has no chromosomal counterpart in the human genome with which to exchange genetic information during meiotic recombination. Thus, the Y chromosome is more genetically homogenous than the autosomes and is thus extremely useful in genetic clock studies. In this study, they noted that if humans have actually been around for several hundred thousand years or more, according to RAO they should have accumulated 8 to 59 times the amount of mutations that we currently observe in Y chromosomes worldwide. Quite notably, they empirically proved that we can only observe about 4,500 years of mutation accumulation in all of human paternal ancestry, as documented in the record of the human Y chromosome.
Genetics corresponds to linguistics
A variety of studies have analyzed genetic diversity in relation to language for isolated regions of the world such as Europe, India, South and Central America, parts of Africa, etc., but none have done this type of work on a global level.25 The initial efforts had been purely asymmetrical in their approach in comparing DNA with various languages. In other words, some studies had focused purely on the genetic analysis and then used linguistics to interpret the findings, while other research analyzed linguistic data and then attempted to interpret the results with genetics. Therefore, little is known about the development of global human demographics because few studies combined genetics and language data in the same analysis.
A fairly recent secular study of direct interest to the creation science community was reported in 2015. It supports a language-based mechanism for the biblical account of human ethnic group origins, i.e. the confusion of languages and the Babel dispersal shortly after the Flood.25 This study was unique because it combined data from three important fields: language, genetics, and demographics. The researchers analyzed the largest available datasets of distinct sound units in a language, called phonemes, in 2,082 global languages and combined this with genetic profiles based on common DNA variants derived from 246 different people groups. They stated: “On a global scale, both genetic distance and phonemic distance between populations are significantly correlated with geographic distance”; and, “There is a relationship between human dispersal and linguistic variation.” Thus, the data derived from this extensive study confirmed the close tracking of both language and genetics for people groups across the globe, but the revealed migration patterns did not follow evolutionary RAO predictions. The researchers also stated: “However, the geographic distribution of phoneme inventory sizes does not follow the predictions of a serial founder effect [single lineage of descent] during human expansion out of Africa.” In other words, there was no clear dispersal pattern coming out of Africa according to the popular RAO dogma.
This author fully recognizes that languages and genetic histories are complex and often don’t directly correlate and that this data cannot be used to reconstruct a clear global picture of descent from Babel. Nevertheless, these results challenge the evolutionary RAO idea of a single lineage of languages and human populations evolving in Africa and then dispersing from there globally.25 While the researchers believed the genetic data by itself offered marginal support for RAO, they were forced to admit that “genetic and linguistic data show similar signatures of human population dispersal within regions”. These results clearly support the fact that modern people groups have multiple origin starting points.
Genetics correspond to population growth
The Bible presents a variety of historical events and paradigms that allow us to predict population growth and interpret this data within genetics and what we know about the current global population. The three primary biblical events would be the original creation of Adam and Eve about 6,000 years ago, the global Flood which produced a genetic bottleneck whereby the earth was repopulated from Noah’s three sons and their wives about 4,300 years ago, and the tower of Babel, whereby multiple people groups originated via the confusion of languages.26,27
These are major, biblical, and global impacting events. In order to study these and other population growth scenarios, such as the growth of the Hebrew people during the Egyptian captivity, Carter and Hardy developed a sophisticated software program for modelling growth under a wide range of parameters and scenarios.26 Their software took into account population size, minimum child-bearing age (females and males), pregnancy probability, remarriage rates, and mortality. They found that it was trivial to obtain the current world population (in 2015) at about 7 billion people from an original ancestral founding group of Noah’s three sons and their wives. They also affirmed the feasibility of the growth of the Hebrew population during the 430 years of captivity in Egypt, as recorded in detail in the Scriptures.
Building on the population research which vindicated the Bible’s chronologies, Carter and Powell began to research the Creation-Flood-Babel model of human origins in terms of population genetics and demographics.28 They created another population modelling program to examine and model changes in allele frequency. Their model created a hypothetical genome of 100,000 alleles contained in 22 independent chromosome arms, with each arm scaled proportionally to its length in the human genome. In a creation scenario of two individuals, alleles were set to a heterozygous state, representing created diversity. Childbirth was controlled by various population parameters. Recombination events for each chromosome arm in each generation were also varied and the alleles tracked in all individuals. At specified points in the model, bottlenecks could be introduced, such as the one imposed by the global Flood. They demonstrated that a biblical population genetic-growth model can accurately account for the current diversity we see among modern humans. In contrast to evolutionary claims, they showed that there is no reason to reject a literal interpretation of biblical chronology based on the allelic diversity or on the distribution of that diversity. They also showed that the genetic risk factor of diversity loss in the Flood bottleneck was not a problem either. In fact, in a later paper, Carter specifically tackled the question of diversity loss in the Flood.29 He estimated that the Ark passengers designated to repopulate the earth could have carried up to 77% of the available pre-Flood created allelic diversity. Therefore, the majority of the original created allelic diversity could have been carried aboard the Ark and used to fuel the repopulation of the earth.
In regard to mutation-based alleles, Carter followed up his previous research by showing that patriarchal drive (an increase in gamete mutation with an individual male’s age) was not only real, but likely a significant factor in global repopulation post-Flood.30 This was especially pronounced during the first few centuries post-Flood where males, on average, still lived much longer than they do currently. As a result, there would be an increase in mutation-based diversity shortly after the Flood during the initial phases of global population growth. Thus, a biblical model of population growth combined with genetic analyses is fully capable of describing both common (created) variant diversity and mutational diversity within modern humans.
In regard to global population growth, Jeanson has recently taken a novel approach by incorporating demographic population growth into his recent Y-chromosome diversity data.31 His work showed how population growth matches up with lineage splits in his previously published Y-chromosome genetic clock data.24 A biblical timescale fitted well with the last 4,500 years of human population growth, as reflected in the phylogenetic branching events for the lineages in the Y-chromosome trees, rather than just in the data represented at the tips of the tree. Quite notably, the Y-chromosome and population growth data fitted well with specific shifts in population structure in biblical ancient history, such as the global famine described during Joseph’s life as a ruler in Egypt. Such a widespread famine would dramatically have stunted population growth and thus it was detected in Jeanson’s genetic branch analyses.
Summary and conclusion
Based on the biblical chronologies and genealogies in the Hebrew Masoretic text, we know that the global Flood recorded in Genesis occurred about 4,500 years ago. The earth was then repopulated by Noah’s three sons and their wives. Furthermore, at the tower of Babel, the confusion of languages imposed yet another event with genetic consequences. As a result of these biblical historical events, it stands to reason that we should find genetic signatures of these events and timelines in human DNA, population growth, and language. As demonstrated in this article, a copious amount of research has been done by both creationist and secular scientists that vindicates the biblical model of human origins and debunks the RAO.
And before much of this research was done in the creationist community, the famous evolutionary geneticist A.R. Templeton aptly foreshadowed the demise of the RAO interpretation of human origins based on genetic data, when he stated in 2005: “The out-of-Africa replacement hypothesis is strongly rejected by the haplotype tree data [genetic-ancestry analyses], and this is also supported by the fossil and current human data.”32
The new Y-chromosome genetic data brings a huge challenge to the secular science community. While discussing the ramifications of this, Jeanson recently stated:
“I’m anxious to see how the evolutionists try to dismiss this second, independent line of genetic evidence for the young-earth timescale. Not only do they have to explain why the data contradict evolution, they have to also explain why the data are such a tight match with the predictions of Biblical creation. And they have to do it both for DNA inherited through females, and now also for DNA inherited through males.”33
Indeed, the empirical genetics research, especially that done within the creationist community over the past 10 years, is truly a sound dismissal of evolutionary RAO speculation and a huge confirmation for the story of mankind documented in the literal history of the Bible.
References and notes
- Henn, B.M., Steele, T.E., and Weaver, T.D., Clarifying distinct models of modern human origins in Africa, Curr. Opin. Genet. Dev. 53:148–156 | doi:10.1016/j.gde.2018.10.003, 2018. Return to text.
- Carter, R., The neutral model of evolution and recent African origins, J. Creation 23(1):70–77, 2009. Return to text.
- Wolf, A.B. and Akey, J.M., Outstanding questions in the study of archaic hominin admixture, PLoS Genet. 14:e1007349, 2018 | doi:10.1371/journal.pgen.1007349. Return to text.
- Hublin, J.J. et al., New fossils from Jebel Irhoud, Morocco and the pan-African origin of homo sapiens, Nature 546:289–292, doi:10.1038/nature22336, 2017. Return to text.
- Richter, D. et al., The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age, Nature 546:293–296, 2017 | doi:10.1038/nature22335. Return to text.
- Thomas, B. and Tomkins, J., How reliable are genomes from ancient DNA?, J. Creation 28:92–98, 2014. Return to text.
- Prufer, K. et al., The complete genome sequence of a neanderthal from the Altai Mountains, Nature 505:43–49, 2014 | doi:10.1038/nature12886. Return to text.
- Posth, C. et al., Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into neanderthals, Nat. Commun. 8:16046, 2017 | doi:10.1038/ncomms16046. Return to text.
- Langley, S.A. et al., Haplotypes spanning centromeric regions reveal persistence of large blocks of archaic DNA, Elife 8, 2019 | doi:10.7554/eLife.42989. Return to text.
- Chen, L. et al., Identifying and interpreting apparent neanderthal ancestry in African individuals, Cell 180:1–11, 2020. Return to text.
- Tomkins, J. and Bergman, J., Evolutionary molecular genetic clocks—a perpetual exercise in futility and failure, J. Creation 29(2):25–35, 2015. Return to text.
- Tomkins, J., Empirical genetic clocks give biblical timelines, J. Creation 29:3–5, 2015. Return to text.
- Parsons, T.J. et al., A high observed substitution rate in the human mitochondrial DNA control region, Nat Genet 15:363–368, 1997 | doi:10.1038/ng0497–363. Return to text.
- Gibbons, A., Calibrating the mitochondrial clock, Science 279:28–29, 1998. Return to text.
- Tennessen, J.A. et al., Evolution and functional impact of rare coding variation from deep sequencing of human exomes, Science 337:64–69, 2012 | doi:10.1126/science.1219240. Return to text.
- Fu, W. et al., Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants, Nature 493:216–220, 2013 | doi:10.1038/nature11690. Return to text.
- Jeanson, N.T., Recent, functionally diverse origin for mitochondrial genes from ~2700 metazoan species, ARJ 6:467–501, 2013. Return to text.
- Carter, R.W., Mitochondrial diversity within modern human populations, Nucleic Acids Res. 35:3039–3045, 2007 | doi:10.1093/nar/gkm207. Return to text.
- Sanford, J. and Carter, R.W., In light of genetics… Adam, Eve, and the creation/fall, Christian Apologetics J. 12:51–98, 2014. Return to text.
- Carter, R.W., Lee, S.E., and Sanford, J.C.; in: Whitmore, J.H. (Ed.), Proceedings of the Eighth International Conference on Creationism, Creation Science Fellowship. Return to text.
- Jeanson, N.T., A young-earth creation human mitochondrial DNA “clock”: whole mitochondrial genome mutation rate confirms d-loop results, ARJ 8:375–378, 2015. Return to text.
- Jeanson, N.T., On the origin of human mitochondrial DNA differences, new generation time data both suggest a unified young-earth creation model and challenge the evolutionary out-of-Africa model, ARJ 9:123–130, 2016. Return to text.
- Jeanson, N.T., Mitochondrial DNA clocks imply linear speciation rates within “kinds”, ARJ 8:273–304, 2015. Return to text.
- Jeanson, N.T. and Holland, A. D., Evidence for a human Y chromosome molecular clock: pedigree-based mutation rates suggest a 4,500-year history for human paternal inheritance, ARJ 12:393–404, 2019. Return to text.
- Creanza, N. et al., A comparison of worldwide phonemic and genetic variation in human populations, PNAS 112:1265–1272, 2015 | doi:10.1073/pnas.1424033112. Return to text.
- Carter, R. and Hardy, C., Modelling biblical human population growth, J. Creation 29(1):72–79, 2015. Return to text.
- Hardy, C. and Carter, R., The biblical minimum and maximum age of the earth, J. Creation 28(2):89–96, 2014. Return to text.
- Carter, R. and Powell, M., The genetic effects of the population bottleneck associated with the Genesis Flood, J. Creation 30(2):102–111, 2016. Return to text.
- Carter, R., Effective population sizes and loss of diversity during the Flood bottleneck, J. Creation 32(2):124–127, 2018. Return to text.
- Carter, R., Patriarchal drive in the early post-Flood population, J. Creation 33(1):110–118, 2019. Return to text.
- Jeanson, N.T., Testing the predictions of the young-earth Y chromosome molecular clock: population growth curves confirm the recent origin of human Y chromosome differences, ARJ 12:393–404, 2019. Return to text.
- Templeton, A.R., Haplotype trees and modern human origins, Am. J. Phys. Anthropol. Suppl 41:33–59, 2005 | doi:10.1002/ajpa.20351. Return to text.
- Tomkins, J.P., Y-chromosome study confirms Genesis Flood timeline, icr.org/article/11732/, 2019. Return to text.


Readers’ comments
Comments are automatically closed 14 days after publication.